Common Dusky-blue
Erina hyacinthina hyacinthina (Semper)
Erina hyacinthina form josephina (Harris)
Western Dusky-blue
Erina hyacinthina form simplexa (Tepper)

Western Dusky-blue
Interesting Aspects
This butterfly is often placed in the all-encompassing genus Candalides but adult and immature stage morphologies, and larval hostplant preference, indicate it belongs in the more compact genus Erina. We have updated the site accordingly. Please report ongoing issues to web@butterflyconservationsa.net.au.
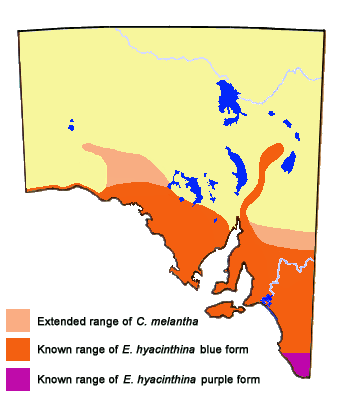
This is a very interesting butterfly, in which the adults occur in several very distinct colour forms. The nominotypical form occurs in the high rainfall coastal and Great Dividing Range areas of southeast Qld, NSW and Victoria. In this form, both males and females have dull purple coloured areas on the wing uppersides. The form simplexa (Tepper) occurs in dry temperate areas of southern Australia, and is present in all the southern mainland states. In this form, both males and females have bright metallic blue coloured areas on the wing uppersides. Where these two forms meet geographically, there is another adult colour form, which can have a mix of purple (violet) and blue on the wing uppersides. This form is called josephina (Harris). In the high rainfall southwest corner of Western Australia, there is another dull purple coloured adult form that is very similar to the nominotypical form. This form is called gilesi (M. Williams and Bollam).
The butterflies normally fly very close to the hostplants, but males also have a strong preference for hill topping, especially when females are scarce. Males set up territories on dune-tops and hilltops, from where they make frequent forays, either to other parts of the hill/dune top or to adjacent hostplant areas to look for unfertilised females, or to fend off other males, but always returning to their territories. They usually perch on twigs or bushes in these territories with wings either open or closed, usually about head-height from the ground. On cold days they will rest on the ground where it is warmer. Females are usually seen flying near the hostplants where they are constantly searching for a good place to lay eggs. Flight also normally occurs at about head-height, and is fast in males, but slow in the females when they looking to lay eggs. If disturbed, the flight is very fast and direct.

Upperside of male form josephina
The wing underside colour is a dull cryptic grey, which provides some camouflage when settled, but is probably more useful as camouflage when in flight. The underside is similar to both the related Erina acasta and Cyprotides cyprotus, and all three can fly together in mallee and dry heathland habitat. The former butterfly differs by being smaller and has a dark tornal patch on the hindwing underside, while the latter has a more brownish underside that lacks the large black subtornal spots in the forewing. In a dune-topping situation, the latter two butterflies usually settle lower than E. hyacinthina. The male blue form of E. hyacinthina is sometimes very obvious in flight due to the bright metallic blue colour on the uppersides of its wings, and a large specimen can be confused with Ogyris amaryllis when it first takes flight from a hidden resting position. The extent of the blue and purple colour on the wings is usually stable, although the blue extent in simplexa females from the hot and dry, northern inland areas of Eyre Peninsula can be more extensive than specimens from the coast or eastern parts of South Australia. Worn specimens of the blue form can sometimes acquire a purplish hue, particularly those occurring proximal to the purple form.
Both sexes can usually be approached with ease when settled.
The divergent colour forms of the Dusky-blues are an example of where speciation is not complete, and where two 'subspecific' populations have come together. The forms of hyacinthina are usually treated as subspecies, although the conventional understanding of the geographic subspecies concept does not easily fit the population taxonomy of the 'hyacinthina' complex. The nominotypical form is likely to be the original colour form that developed in the evolutionary ancient, temperate forests that were once widespread through much of southern Australia. As the interior of Australia subsequently dried out to produce the mallee woodlands, the blue form of the butterfly developed in unison with this dry woodland along with its arid tolerant Cassytha melantha (Lauraceae) larval hostplant. At some evolutionary recent stage there was probably geographical isolation of the two colour forms, but which was not long enough for complete speciation. Both blue and purple colour forms use different Cassytha species for larval hostplants. However, in captivity the larvae of the Common Dusky-blue can eat and survive on the Cassytha used by the Western Dusky-blue, whereas the natural Cassytha hostplants of the Common Dusky-blue are toxic to the first instar larvae of the Western Dusky-blue. In South Australia (and north-west Victoria) at least, this suggests that where the two colour forms meet at the limits of the Cassytha melantha, interbreeding occurs. It is probable the gene(s) controlling the blue upperside wing colour of the Western Dusky-blue is dominant over the purple colour as the blue and purple forms are very rarely seen flying together, and then only at their contact zone. The purple form has never been seen flying with the blue form within the main flight zone of the blue form. The fertile mixed 'hybrid' colour form josephina likely represents the regressive portion of the progeny that has retained most of the attributes of the purple form. The early stages of the latter butterfly behave like the earlystages of the nominotypical form. The hybrid form has a limited range and quickly disappears as it interbreeds with the nominotypical form. Hence the blue and purple colour forms cannot, in a general sense, occur together, which has caused the distinct distributions of the two forms. Some scientists would suggest that since the blue and purple colour forms are incompatible in the 'wild', then they might be best treated as separate species. DNA techniques may provide a resolution, although a laboratory-interbreeding program would be more definitive. (A similar relationship occurs between the Lycaenid species Jalmenus eubulus and Jalmenus evagoras in Queensland that was interpreted on the basis of DNA studies, Eastwood et al. 2008, wherein it was concluded that the two should be treated as full species). A similar contact of 'hyacinthina' butterflies occurs in southwest WA where purple form gilesi feeding on Cassytha racemosa meets blue form simplexa feeding on C. melantha and C. aurea. C. racemosa can also occur with the latter Cassytha, but the opposing ranges of the two butterflies cease at the range limits of the C. melantha and C. aurea growth. In central and northern NSW another closely related purple species Candalides geminus (feeding on C. pubescens) separates form simplexa from nominotypical hyacinthina, possibly suggesting a more advanced previous contact between the latter two forms.