

Male 39 mm from Eyre Peninsula east
Female 47 mm from Eyre Peninsula central
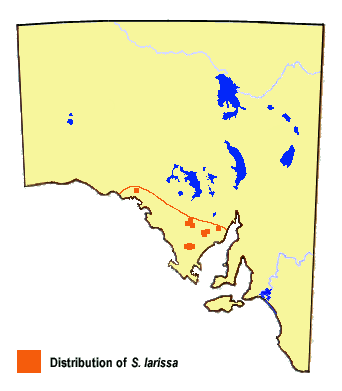
A common species, restricted to temperate areas of Eyre Peninsula. Its primary larval hostplant is Lepidosperma carphoides (Black Rapier-sedge) (Cyperaceae), which is found only on the east side of the peninsula. In the latter area and in other areas where the sun-moth flies in the absence of L. carphoides, females are attracted to Lepidosperma congestum (Clustered Sword-sedge) and Schoenus racemosus (Tufted Bog-sedge) (Cyperaceae), which are also likely hostplants.
Synemon larissa belongs in a similar cryptic-patterned group of sedge feeding sun-moths in South Australia that include S. discalis and S. parthenoides, and possibly other as yet unstudied species from West Australia. The group adults also possess a functional proboscis, and in combination with their different genitalia and other morphological and feeding attributes, suggest they belong in their own genus as compared to the grass feeding S. collecta group. S. larissa differs from S. discalis mainly by its larger size, different male genitalia, and several small differences in wing morphologies, such as the upperside (UPS) ground colour of the forewings (FW) is black in S. larissa (SL), but brown-black in S. discalis (SD); there is not a narrow central black transverse zig-zag line in the central part of the FW UPS as in SD; the inner margin (next to abdomen) orange coloured area of the hindwing (HW) UPS is joined to the central HW orange spots in SL, but usually not in SD.
S. larissa differs from S. parthenoides (SP) by different male genitalia; the antennae clubs in SL are brown-black, but brown in S. parthenoides; the UNS HW costal spots (next to the FW) in SL are white coloured, but only weakly so or absent in the typical SP although similar to S. parthenoides valma; the large black spot in the UNS FW area next to the costa (the leading edge of the wing) in the basal part of the wing closer to the body is closed off in SL to form a complete circle, but is open in SP. S. larissa also has a morphology and size similar to S. selene but the 'ground' colour in the latter is brown rather than black, and the former tends to fly in spring rather than late summer-early autumn as for S. selene. Interestingly, S. larissa has morphological attributes comparable to both S. discalis and S. parthenoides, and in combination with its present geographic isolation might suggest that S. larissa originated as a hybrid of the other two species occurring in the past during the late Pleistocene climate cycling event (last ice age ~15,000-70,000 years ago). S. larissa replaces S. parthenoides on Eyre Peninsula.
It has a habitat preference for mallee woodland in which the larva hostplants grow. Males have a wing expanse of 34-42 mm and females an expanse of 46-52 mm, and tend to fly in spring during October-early November. Flight starts later than the flight of S. discalis if they occur together. They fly near ground level or just above the top of their low growing hostplants. Typical of most sun-moths, the males start to fly earlier in the season than females, and the flight season for both sexes tends to be earlier in the northern parts of its range. It is active during most of the warm sunny periods of the day. Males become active early in the day, first feeding from flowers then tend to fly about the hostplant habitat looking for females to mate with, particularly those that have newly emerged from their chrysalis. Eventually the males settle down, often setting up small leks in open or cleared areas like vehicle tracks, waiting for females to fly past. If the habitat is suitable, they will also set up lek (courtship) territories on the tops of dunes, ridges and hills near the hostplants. They will either perch on low dead twigs or rest directly on the ground. Periodically, if no females or other disturbances occur they will fly off to patrol the hostplant habitat looking for females, and if unsuccessful will not necessarily return to the same lek. After mating the females once more become active, laying eggs until late afternoon, when they start to feed from flowers before eventually seeking a place to spend the night.
This species may locally have mass emergences of both males and females when hundreds of them may occur together, causing incessant territorial battles and mating rituals for the males.
The female lays its eggs below ground adjacent to or on the stems of the hostplant. This she does by either walking up to the plant on the ground if possible, or by working her way down within the plant from the top. She uses her very long stiff ovipositor, that can be extended from the end of her abdomen, which she probes down the sides of the hostplant leaves and then lays the single egg as deep as she can place it. The eggs are normally pale yellow when newly laid but eventually turn white near larval eclosion. They are ellipsoidal spindle shaped about 2.1-2.9x0.85-1.0 mm and have 8-11 prominent longitudinal ridges, linked by many (~50) finer cross striae. The first instar larva ecloses (emerges) after about 19-32 days, is 3.0-4.5 mm long and subtranslucent pale yellow coloured with a yellowish brown head. They likely live below ground within the culm and root zone of the hostplant similar to the other members of the group. Its mature larvae and pupae are undescribed.
The adults are often seen nectaring on flowers such as the white flowered Homoranthus wilhelmii (Eastern Feather-flower), Calytrix sp (Fringe-myrtle) and a yellow flowered Glischrocaryon behrii (Golden Pennant). S. larissa is not presently threatened. The type male came from east Eyre Peninsula.



