-
Larval Food Host
-
Subspecies idothea utilises the saw-sedges
Gahnia clarkei (tall saw-sedge or cutting grass) and
G. radula (thatch saw-sedge), and also very rarely
G. trifida (cutting grass).
Subspecies clara utilises G. sieberiana (red-fruit saw-sedge)
and G. trifida (Cyperaceae).
G. seiberiana is preferred as a hostplant ahead of G. trifida.
Interestingly, the former hostplant does not occur naturally in the Southeast Region.
The larvae eat the leaves of the hostplant. The skipper normally requires its hostplant
to be in the shade.
-
Eggs
-
Large (1.5 mm along the long axis), hemi-ellipsoid (nearly hemispherical) with a flat
base, and with about 23-28 fine vertical ribs, and the micropylar area on top of the
egg is depressed. Pale green when new, but after 2 days if fertile the eggs become
pale yellowish green and acquire a ragged red lateral ring and a similarly coloured
micropylar area or circular dorsal patch. Laid singly on the leaves of the hostplant,
usually on the lower parts of the plant on the underside of a leaf near its edge.
The egg shell is eaten by the larva after its emergence.
-
Larvae
-
The first instar is long cylindrical, the forward half of the larva is pale
greenish yellow and the posterior half of the larva is pale yellow with five
narrow longitudinal dorsal and subdorsal brown lines that do not continue onto
the anal plate. There is a large shining black head having a few long hairs,
the neck (prothoracic plate) is dark brown, the neck area is dark wine red,
and long recurved hairs occur posteriorly. After eating the empty eggshell the
larva will eat a small portion of the hostplant to test that it is the correct
hostplant, then it moves to near the tips of the leaves to form a small tubular
shelter opening at the top, by silking together two or three leaves. The young
larva gradually turns green as it eats the hostplant.
Subsequent instars gradually acquire indistinct pale and dark longitudinal lines
and lose the long posterior hairs, and the head becomes brown and eventually by
about the fourth instar, acquires the pale brown head pattern described for the
final (fifth) instar larva. New silk lined tubular shelters are periodically
constructed to fit the growing larvae, using leaves of the hostplant, the internal
diameter of the shelters being a close fit to the diameter of the larvae.
The shelters are sealed at the base and open at the top (towards the tips of the
leaves), and in the case of this skipper, the leaves are usually silked together
in a straight manner. If the shelter is made in the outer parts of the hostplant
then during its development the larva will eat the leaves back from the tips
towards the shelter entrance. However, the late stage instar larvae of this skipper
more often will utilise the tubular base of a single leaf that has separated from
the main part of the tussock base, which gives the larva a ready-made tubular
shelter. These larvae will then eat the leaf edges from anywhere along the length
of the long leaf, or will eat down from the top an individual leaf leaving a
characteristic oblique edge across the end of the leaf. Larvae feed at night for a
very brief period (usually very late at night), hiding from predators during the day
inside their shelters.
The final instar is long cylindrical, about 35-40 mm long, with the last posterior
segment flattened into the anal plate, smooth, but with the posterior end having some
hairs. Green coloured, sometimes bluish or yellowish, with a darker dorsal line, and
sometimes there are other indistinct longitudinal lines when the larva is still in its
early final instar growth. The head is large, rugose, elongate and slightly flattened,
there is a distinct central longitudinal furrow, the top is rounded, with a few long
hairs ventrally and there are also some very short frontal hairs that are bent and
directed forwards. Pale brown coloured with a moderately broad, brown longitudinal
median line in the front that expands ventrally and tapers to a point at the dorsal
apex, the mouth parts are dark brown and there is an inverted pale brown coloured V
mark immediately above the mouth, and there is also a yellow patch on either side of
the mouth parts. The side of the head has a narrow brown vertical stripe, which continues
to the top of the head and joins with the brown frontal triangular mark. The body, and
particularly the anal plate on the last segment, is covered in minute secondary setae
that are wine glass or vase shaped, and which are set on simple smooth raised bases that
are transparent along the body but brown coloured on the anal plate. Near pupation the
larvae turn pale semi-translucent green and have a dark longitudinal dorsal line.
The final larva shelter is constructed either in the outer half of the plant, or at the
base of an individual leaf, and is up to 7 cm long. Shelters formed in the outer parts
of the hostplant are made by joining up to six hostplant leaves together, to form a tight,
strongly silk lined, straight tubular structure, sealed at the base and opening at the top.
Larvae development is usually rapid in summer and autumn, and they continue to eat and
develop very slowly through winter, and pupation occurs in the following spring or summer.
The presence of larvae for this species (and for all other Hesperilla larvae) on
the hostplant are discernible by either the distinctive looping of the hostplant leaves
resulting from the construction of the larval shelter in the outer parts of the hostplant,
or by large serrated eat marks along the edges of the leaves, or by the leaf above the
shelter being truncated at an oblique angle. The leaves used in the construction of the
shelter are tightly fixed by silk to form the shelter, and as the leaves continue to grow
(from the base) the unequal growth rate of each leaf causes the fastest growing leaves to
produce a loop beneath the shelter.
-
Pupae
-
Long cylindrical, about 23-27 mm long, pale whitish green to pale green,
sometimes yellowish or brownish, covered in a moderately heavy white
powdery bloom, the latter acting as a water repellent and perhaps fungicide.
Some brown coloured, very short posteriorly directed spinose bristles on the
abdomen, which become more abundant posteriorly, and which arise from conical
bases. The posterior end tapers to a brown, rugose, dorsally ridged cremaster,
that is flattened, and strongly divided and expanded into a bifid shape similar
to 'Mickey Mouse' ears. The head is rounded, with the head cap (operculum) being
black and brown coloured, having a characteristic rounded shape, and there are
some short hairs that are pale coloured and spinose. The central part of the
operculum is black coloured, heavily sclerotised (very rugose) and is divided
into three, distinctly separated, and strongly elevated (anteriorly projected)
parts. A larger oval shaped ventral part, and two smaller equal-sized, circular
dorsal parts. There are further, smaller less sclerotised brown coloured areas
lateral to the central part.
Pupation occurs in the final larval shelter on the hostplant, and the larva pupates
head upwards towards the leaf tips. The pupa is secured within the silk lined shelter
by very strong hooked bristles emanating from the terminal-ventral parts of the
cremaster, (same principal as velcro). The pupal duration is about 22 days in late
spring for subspecies clara near Mt Lofty, while for subspecies idothea
it is about 13-20 days in late summer. The empty pupa case remains inside the shelter
after the adult skipper emerges, and is nearly transparent in a very pale brown or
whitish colour.
The shape of the operculum and cremaster, the colour of the pupa (alive or as an empty
case), and the morphological properties of the larva or discarded larval skin are
diagnostic for the species and can be used during field surveys to differentiate
H. idothea from other Hesperilla species in South Australia outside of
the flight times for the adult skippers.
-
Flight Period in South Australia
-
It is single brooded. Subspecies clara flies from November to January,
while in the Lower Southeast, subspecies idothea has an extended flight
from December to March. In the nearby Grampians Region of Victoria the latter
subspecies starts to fly in late October.

-
Distribution
-
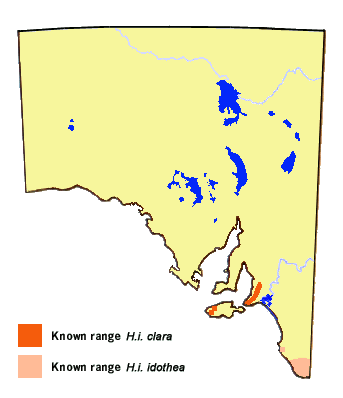
Subspecies clara is only found along the South Mt Lofty Ranges and at
the western end of Kangaroo Island. Within South Australia, the nominotypical
subspecies idothea is restricted to the Lower Southeast Region.
The latter also occurs in the cool moist areas of Victoria and along the Great
Dividing Range in NSW and southeast Qld, and it also occurs in Tasmania.
The range of subspecies clara has been in a steady decline along the South Mt
Lofty Ranges due to urbanisation and degradation of creeklines, and now can only be
found near to Mt Lofty or the extreme south of the Fleurieu Peninsula.
-
Habitat
-
This skipper is essentially a forest skipper, preferring the cool recesses of
the forest, but it will venture into cool, moist open woodland. It requires
pristine habitat and needs a cool microclimate of shady trees. Subspecies
clara is confined to saw-sedge bearing shady creeklines and upland
spring environments in cool-temperate, high rainfall elevated areas of the
South Mt Lofty Ranges, and western Kangaroo Island. Subspecies idothea
is found either in Gahnia radula bearing forests, or in open
Gahnia clarkei wetlands that are surrounded by a shady cover of native
trees and large bushes.
-
Conservation Status in South Australia
-
The mainland habitat of both subspecies in South Australia is now severely
fragmented and degraded, due to urbanisation and agricultural use.
The habitat of subspecies clara has been the worst affected and its
status is vulnerable on the mainland. On Kangaroo Island where there has been
a better retention of wetland habitat the butterfly is believed to be still
stable, but very localized in distribution. The nominotypical subspecies is
more stable due to the presence of its habitat in conserved native forests
and conservation parks, and can be considered uncommon. This subspecies can
be locally common in the extensive remnant forest areas of the eastern states.
-
Threats
-
This skipper has suffered considerably from the agricultural and urban disturbances
of its saw-sedge wetland habitat. Remnant saw-sedge bearing creeklines and wetlands
continue to be drained, dammed and degraded. Most creeklines are now choked and
smothered with weeds and introduced trees, particularly African feather-grass, ash,
blackberry, broom, couch, ivy and kikuyu. Irresponsible disposal of urban and
agricultural waste products and toxins inevitably end up in wetlands contributing
to their degradation.
-
Conservation Strategy
-
The retention of pristine saw-sedge wetland habitat is essential for the survival
of this skipper in South Australia, and specific habitat (particularly for subspecies
clara) may have to be conserved for its long-term survival. On the mainland
there needs to be a major revegetation of suitable creeklines with the saw-sedge
hostplants, and the latter should be included in all suitable revegetation projects.
Remaining saw-sedge wetlands used for pastoral purposes need to be managed in an
ecologically sustainable manner, and ideally, suitable wetlands need to be fenced
off from the destructive feeding habits of grazing domestic stock. (Unfortunately,
most wetlands on private land, usually dairy-land, are used as a backup food source
for the cattle due to the adjacent pasture land having been severely overgrazed.)
The control of mosquitoes in remaining broad-acre saw-sedge wetlands through the
use of broad spectrum insecticides by aerial means needs to be judicially controlled,
but preferably should cease, as it can be a major cause of fauna destruction.
The draining of remaining pristine wetlands needs to cease, as this practice causes
summer stress to the Gahnia that is ultimately fatal to Hesperilla
larvae. The dumping of urban vegetation waste into creeklines and wetlands needs to
cease. There needs to be a major public education process about the continuing
degradation effects on wetlands.






