-
Larval Food Host
-
Neurachne tenuifolia (mulga-grass) (Poaceae).
This grass has a tussock habit with low, basal involute filiform leaves,
but long erect flower stalks. The larvae eat the leaves of the hostplant.
-
Eggs
-
Small, slightly hemi-ellipsoid, 21-29 prominent vertical ribs, with indistinct
cross ridges, the small micropylar area on top of the egg is depressed. Pale
yellow when newly laid, and the eggs do not later acquire red markings if
fertile. Laid singly on the leaves of the hostplant, although with successive
layings (by the same or different females) more eggs may occur on a single
blade of grass. Up to 25 eggs have been recorded on a single plant. Green or
dead leaves are used for laying, although the former are preferred. Eggs are
laid on the convex surface of the leaf, in the outer half, and the base of
the egg will wrap itself around the stalk during the hardening stage of the
egg, similar to that of its sister species
Herimosa albovenata.
Larvae start to hatch (eclose) after 8 days. The egg shell is eaten by the
larva after its emergence.
-
Larvae
-
The first instar is long cylindrical, initially entirely pale yellow
but gradually turn green after eating the hostplant, with a large
pitted very dark brown head having some long hairs, the neck
(prothoracic plate) is brown, and long recurved hairs occur posteriorly.
After eating the empty egg shell the larva will move basally to the
cavity area of the ligule-sheath attachment area of the leaf to the
stalk, nearer the base of the plant to make its initial shelter.
The young larva rests in the shelter during the day with its head
pointing upwards towards the leaf tip. The larva eats nocturnally
by feeding on the edges of the leaf.
Subsequent instars are similar to C. arenaria and gradually
acquire pale and dark longitudinal lines and lose the long posterior
hairs, the head gradually becomes paler and gets a brown frontal
marking and eventually by about the fourth instar acquires the head
pattern for the final (fifth) instar larva. New shelters are periodically
constructed to fit the growing larvae, by using the basal parts of
the leaves and stalks of the hostplant, and the internal diameter of
the shelters is almost an exact fit to the diameter of the larvae.
The shelters are open at the top (towards the leaf tips) and sealed
with silk at the base, which is unique for this tribe of skippers.
Larvae emerge from the shelters to eat leaves at night.
The final instar is long cylindrical shaped, about 16-22 mm long,
and very similar to C. arenaria.
This skipper has developed survival means to overcome the periodic
extended dry periods of Central Australia, and which is another
feature common to the tribe of skippers in which this skipper is
a member. Its mature larvae can enter into a dormant diapause
condition when the hostplant grass becomes too dry or rank to eat.
These larvae can diapause during the dryer months until the rain
occurs. They then break diapause to feed slowly on the rejuvenated
green growth of their hostplants until their pupation.
The presence of larvae on the hostplant is discernible by the
characteristic long (up to 5 cm long), shallow, jagged sawtooth-like
edges to the leaves.
-
Pupae
-
Similar to C. arenaria about 16 mm long. Pupation occurs in the
final larval shelter on the hostplant, and the larva pupates head
upwards, directed towards the leaf tips. The pupa is secured within
the silk lined shelter by very strong hooked bristles emanating from
the end of the cremaster. The pupal duration has not been documented.
The empty pupal case remains inside the shelter after the adult skipper
emerges, and is brown coloured.
-
Flight Period in South Australia
-
Known to fly from mid September to late April. The fight is likely
opportunistic dependant on the infrequent rains that occur in Central
Australia, but primarily on heavy monsoon thunderstorms during late
spring to autumn that promote rejuvenated growth of the perennial
hostplant grasses. This would lead to a split flight period based
on the winter and monsoonal rain periods. Good winter rains would
promote an early spring flight, while good monsoon rains would
promote a late summer-autumn flight. The brood period has not been
documented. Observations during the hot summer months show that
the early stages of the skipper are present on the hostplants
in all stages of development. The skipper overwinters as larvae.

-
Distribution
-
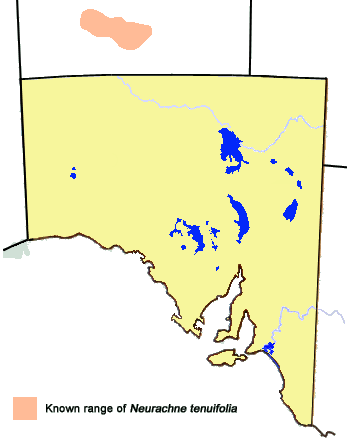
Presently only recorded from Central Australia in rocky hill habitat of the west
MacDonnell Ranges where its foodplant grass Neurachne tenuifolia (mulga-grass)
occurs. However, a similar species Neurachne munroi (dwarf or slender-headed mulga-grass)
also occurs in Central Australia and northern South Australia in similar habitat,
and it is possible that with further research it may be found that the skipper could
also utilise this grass.
-
Habitat
-
Presently, the distribution of the hostplant Neurachne tenuifolia is believed
restricted to Central Australia in rocky hill habitat of the west MacDonnell Ranges.
The skipper tends to fly in the higher parts of the ranges, suggesting it prefers
the cooler parts of its habitat, in particular the south side of the ranges where
the hot sun would have less direct affect on both the skipper and the hostplants.
-
Conservation Status in South Australia
-
Due to sparse records, it was initially believed to be endangered.
With more surveys, new information suggest the skipper is very local
in its status.
-
Threats
-
The main threats for the skipper would be extended drought and the effects
of pastoral disturbances through over-grazing and trampling by animals,
and the occurrence of grass fires. Although the grass can regenerate from
fire provided it is not too frequent, the skipper early stages would be killed.
-
Conservation Strategy
-
The hostplants seem to have a restricted distribution and growth density,
and consequently require protection. Remaining remnant areas of pristine
native vegetation containing the hostplants need to be protected or not
be unecessarily cleared or burnt off. Feral grazing animals would need
to be kept under control.

{kind=link}