-
Larval Food Host
-
Larvae are polyphagous and have been documented to eat numerous hostplants from a variety of
families (presently estimated at 17 families Australia wide). Those hostplants which can be
found in South Australia include Cassinia spp,
Olearia axillaris (coast daisy-bush) (Asteraceae);
Brachyloma daphnoides (daphne heath) (Epacridaceae);
Acacia spp incl. **A. decurrens (queen wattle),
A. leiophylla (smooth-leaf wattle),
A. longifolia longifolia (Sydney golden wattle),
A. mearnsii (black wattle), A. pycnantha (golden wattle),
**A. saligna (golden-wreath wattle) (Mimosaceae);
**Agonis flexuosa, Eucalyptus spp (Myrtaceae);
Banksia sp, Grevillea sp (Proteaceae); Pomaderris sp (Rhamnaceae);
*Prunus sp (plum), ornamental *Rosa sp,
*Rubus "fruticosus" (blackberry) (Rosaceae);
Camellia sp (Theaceae),
Choretrum glomeratum (berry broombrush or common sour-bush),
C. spicatum (spiked sour-bush), Exocarpos aphyllus (stiff cherry),
E. cupressiformis (native cherry) (Santalaceae);
**Alectryon (Heterodendrum) spp,
**Cupaniopsis anacardioides (tuckeroo),
Dodonaea spp incl. D. humilis (dwarf hop-bush),
D. viscosa angustissima (attenuata) (narrow-leaf hop-bush) (Sapindaceae),
Brachychiton sp (Sterculiaceae).
In South Australia the documented hostplants are few. On Eyre Peninsula the hostplant is
Choretrum glomeratum; on Yorke Peninsula the hostplants are Acacia leiophylla,
Choretrum glomeratum, Dodonaea humilis and Exocarpos aphyllus;
near Adelaide they used to use Acacia pycnantha, while in the Upper Southeast
the hostplants have yet to be recorded. In the adjacent parts of western Victoria they have
been recorded on Brachyloma daphnoides and Choretrum spicatum.
The larvae usually eat the leaves and soft stem parts of the hostplants, and eat by scouring
the cuticle surface such that the underlying parts of the plant eventually die and turn brown,
causing the grazing area to look withered and burnt, or in the case of large leaved Acacia
cause the leaves to become skeletonised. If the colony is large then the larvae will even eat
the green bark off the stems and trunk (from either inside or just outside the byre), which may
eventually lead to the death of the hostplant.
-
Larval Attendant Ant
-
In South Australia, larvae are attended only by a small dark-brown ant
Papyrius sp (nitidus group).
It is a byre-building ant with a distinctive coconut-oil smell. The ants themselves
do not live in the byres, which are made of matted fine vegetal debris constructed
by the ants about the base of the larval hostplant to house and protect the larvae
and pupae of the butterfly from predators during the day. These semi-permanent byres
can cover just a portion of the hostplant base, or sometimes can be extensive,
surrounding the entire base of the hostplant at heights of 20 cm or more, sometimes
even extending as narrow extensions up the trunk of the hostplant, and in extreme
cases can spread as branching extensions away from the hostplant for over a metre or
more over decaying logs and other vegetal material on the ground. The ant nest occurs
either deeper underground or at some distance from the hostplant. The gap between the
byre and the trunk of the hostplant is usually not much wider than the thickness of
the mature larvae and pupae. The byres are not always present, but can be quickly
constructed by the ants after eggs are laid by the female butterfly, to accommodate
the subsequent emergent larvae.
As for the Ogyris butterflies there is a symbiotic relationship between the ants
and the early stages of the butterfly. The ants provide accommodation and some protection
for the early stages, while the larvae provide liquid food for the ants from a special
nectary gland (Newcomer's organ) situated at the posterior end of the larvae.
There are several ants that construct byres in SA, some of which can sting, but the
small Papyrius ant is harmless and has a characteristic coconut oil smell that
is irresistible to cats. The compound(s) producing the smell are also found in catnip
and consist of nepetalactone (nepetapyrone) derivatives. It has been found that these
compounds have antibacterial and insecticidal repellent properties (to small wasps
and flies), and so these ants provide further symbiotic service to the H. ignitus
larvae by supplying antibacterial measures inside the byre and also may prevent predatory
small parasitic wasps and flies from attacking the eggs and larvae of the butterfly.
It is not yet known if the nepetalactone compounds can prevent the introduced large
European wasp from destroying the ant byre colonies containing the H. ignitus
larvae. It is also not yet known if the ants produce their own nepetalactone compounds
or whether they sequester them from plants. Several other ants (such as Iridomyrmex
and Crematogaster species) also have these nepetalactone compounds in their
systems, and some of these ants also provide a symbiotic service to other Lycaenid
butterflies.
-
Eggs
-
Small, greyish white, hemispherical shape, basally flattened, the top is weakly domed.
The micropylar area is very small at the egg apex and dark coloured. The sides are
finely reticulated, with mostly hexagonal facets, and some pentagonal adjustment facets.
The facets in the lower half of the egg and again adjacent to the micropylar area are
smaller than those in the upper half. There are raised, tapering, short projections
ending with a hair-like spines, occurring along the facet ridges, over most of the egg
but particularly in the upper half.
Eggs are laid in small batches near the base of the intended hostplant or sometimes
also on vegetal debris at the base of the hostplant. Individual egg batches to 18
have been noted, but the aggregations of several females can lead to much larger
batches. The prime ingredient for egg laying is the presence of the attendant ants,
and although the hostplant choice does not seem to be critical the butterfly does
have a preference in South Australia for broad-leaved Acacia and
Choretrum glomeratum. If an ant byre is not already present, then the ants
will quickly extend or build a new byre to cover the eggs. The eggs begin to hatch
in 10 days. The female butterfly has an egg laying preference for small immature
plants about one metre in height, and if the colony is vigorous and contains many
larvae these plants can be quickly destroyed over a single season.
-
Larvae
-
The first instar is initially pale greyish white with a pale pink dorsal line. Long,
flat onisciform shape, with a slight dorsal keel. The head is very large, smooth,
shiny, with a few short bristly dark setae (hairs), dark brownish black coloured
and partly hidden beneath the body. There is a large dorsal prothoracic plate behind
the head, rectangular shaped, dark greyish black, with two pairs of long forward
directed dark hairs. The posterior dorsal anal plate is grey, small, triangular shaped.
There are very long white (basally darker) peripheral hairs, shorter posteriorly,
two per segment laterally, but with additional hairs anteriorly and posteriorly.
There are dark paired hairs dorsally; on the 2nd segment there are two widely spaced
long recurved hairs, the third segment with two pairs closely spaced shorter hairs,
segments 4-6 with one pair of shorter slightly recurved hairs, segments 7-9 with one
pair of very short knobbed bristly hairs. The dorsal posterior nectary organ is well
developed. The posterior dorso-lateral organs are not developed, but are replaced by
a pair of specialised flattened white moveable short flap-like setae. A further double
pair of similar but longer specialised setae situate between the previous short
flap-hairs and the anal plate, and which extend over the latter plate. There are a few
tiny dark secondary setae along the body, elongated vase-shaped. The initial body length
is about 1.8 mm, but including the long hairs is about 3.1 mm. After eating, the first
instar gradually becomes yellowish pink coloured, with a broad dark dorsal line, a yellow
subdorsal line and several dark narrow lateral lines. The second instar acquires the
posterior dorso-lateral organs, while by the third instar the flap-like setae adjacent
to the latter have completely been lost.
Intermediate instars between the first and final instars are onisciform (slater shaped)
and gradually lose the long dorsal setae, and also gradually become mostly brown coloured
and acquire the pattern of the final instar. When the larvae undertake moulting the head
and prothoracic plate separate joined together, but separately from the rest of the body
skin.
First instar larvae are reported to sometimes remain exposed on the leaves of the hostplant.
Subsequent instars at least, hide during the day, and feed at night. They shelter during
the day inside the ant byre at the base of the hostplant, or deeper still along chambers
constructed by the ants around the base of the hostplant or along its roots. As many as
30 larvae have been reported to live inside these byres. Sometimes if the colony is large
they will also hide in curled leaves higher on the hostplant that the ants have converted
into shelters by sealing off the exposed parts with byre material. At night the larvae
leave the byre, along with numerous ants, to feed on the hostplant leaves. The larvae will
feed gregariously together. The presence of larvae on a hostplant is readily discernible
by the presence of the ant byre, and by the scoring and skeletonising of the hostplant
leaves.
The final instar can either be a fifth or sixth instar stage, depending on how quickly the
earlier stages evolved, and the time of year. When fully mature it is about 14-18 mm long,
elongate onisciform shaped, the lateral edges are scalloped, and the anterior and posterior
areas are flattened. There are some short peripheral hairs that are longer anteriorly and
posteriorly, and also some very short dorsal hairs. Generally greyish, pinkish or yellowish
brown coloured,with a pale narrow longitudinal dorsal line that is continuous onto the
prothoracic plate, pale and dark dorsal and subdorsal chevron markings, and a pale yellow
longitudinal and hachured subdorsal line, edged dorsally dark brown. The prothoracic and
anal plates are large, mostly black coloured, smooth but densely covered in secondary setae.
The body is densely covered in these tiny secondary setae, which consist of an open cup on a
short stalk set on a raised simple smooth base. The cup is variable, either symmetrical or
asymetrical, with the rim being either smooth, crenulate or with about 5-6 short lateral
points and sometimes a central short point. The secondary setae impart a scabrous appearance
to the larvae. The posterior dorso-lateral organs are well developed. The head is small,
smooth, dark brown or black, hidden beneath the body.
The larval period is variable, depending on the location, time of the year, and the condition
of the hostplant, but development is generally slow. During winter, larval development is very
slow. Over the warmer months the colonies usually contain larvae at all stages of development.
-
Pupae
-
About 9-13 mm long, greyish brown, speckled with darker brown markings, long cylindrical,
the abdomen is slightly arched, smooth, rounded anteriorly, the posterior end is also
rounded but is noticeably extended into a pad that terminates beneath with the cremaster.
Pupae are attached to the hostplant by anal hooks and a central girdle. Pupation occurs
either within the byre or more often underground in ant chambers about the base or roots
of the hostplant. The latter is preferred as the larvae are cannibalistic and will eat
other pupating larvae, and there is a better chance for a successful pupation in areas
distal from the byre. The larvae will pupate gregariously together. The empty pupal
skins are eventually destroyed by the ants or by other pupating larvae. The pupal
duration is about one month. The emerging butterflies leave the chambers and byre to
expand their wings.
-
Flight Period in South Australia
-
The butterfly flies throughout the warmer months, with records from late October to
early May. The main flight is late spring to early summer, with a later secondary
flight in late summer and early autumn. Butterflies flying after mid-summer produce
larvae that over-winter.

-
Distribution
-
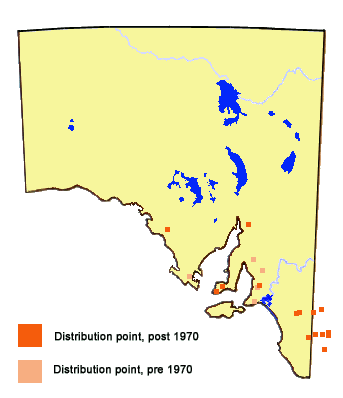
The butterfly used to occur in most areas of the southern agricultural belt having
average annual rainfall in excess of about 300 mm. It occurs in discrete colonies,
which in SA are usually very limited in size, although the population in the Upper
Southeast is quite large. It has not been seen on Kangaroo Island. There used to be
one colony in a conservation park in the Adelaide Hills where it had been breeding
continuously on Acacia pycnantha over an area of about one hectare for 40
years. Sadly this colony became defunct in the mid-1970's. The local nominotypical
subspecies of the butterfly also occurs in similar higher rainfall woodland habitat
in the eastern mainland states, usually within 200 km of the coast. Four other
similar subspecies occur in southwest WA, northern Australia and southern Papua
New Guinea.
-
Habitat
-
The butterfly can occur in many habitats, from dry mallee woodlands and heathlands
to moist forests, but they must be in pristine condition. The colonies can sometimes
occur in roadside reserves, but they are always near large areas of pristine habitat.
It is not found in arid or cold alpine areas. Hilly or dune terrain is preferred,
although not essential. The early stages are totally dependent on only the one species
of attendant ant.
-
Conservation Status in South Australia
-
The butterfly is widely distributed but threatened and considered to be vulnerable
to any interference. Colonies appear to be stable in the large conservation parks
in the Upper Southeast Region, but elsewhere they are very unstable. The butterfly
is more common in NSW and Qld.
-
Threats
-
This butterfly is unable to adjust to agricultural and urban disturbances. Butterfly
collectors frequently target it, and the manner of collection is often terminal for
the colony. Hilltops are usually essential for males as lekking areas, and in urban
fringe areas these are often used for housing and communication towers, and the
butterflies are therefore likely to suffer. Its main threat in conservation areas is
likely to be bushfires, although recent evidence suggests some colonies can survive
'cool fires' if they occur beneath the ground along the roots of the host plants.
However to subsequently exist, the larvae would have to either become cannabilistic
or eat the bark off the roots of the hostplant while it is still moist. Whether the
pupae then undergo long diapause until hostplant regrowth occurs is not known. It is
also not known if the larvae can temporarily adapt to other less preferred hostplants
(of which there are many) while its preferred hostplants regenerate after fire. It is
totally dependent on the one species of ant, which is itself quite rare, and usually
dependent on undisturbed habitat. Burrowing animals such as rabbits, bandicoots and
echidnas (and probably also mice during their periodic plagues) have the potential to
destroy larval colonies at the base of the host trees. A proliferation of bird life
is also a potential threat to the butterfly, as its dependence on hill topping places
it in a situation where it becomes vulnerable to repetitive bird attacks. Perched male
butterflies often display evidence of bird attacks to their wings.
'Friends of native vegetation' and 'Bushcare Volunteers' may also mistakenly destroy the
byres and larval colonies, thinking them to be affecting the vegetation (which they are),
but the plants they affect are much more common than the butterflies. (The ant byre should
not be confused with byres produced by borer beetles and moths, as these byres are very
small in area and occur higher up on the plant.) Opening up the byre exposes the early
stage larvae and pupae to predators, particularly parasitoids, wasps and other ants, which
will quickly destroy the colony. The attendant Papyrius ants cannot fully protect
the early stages once the byre has been opened.
As the byred colony exists above ground, and due to the proliferation of the introduced
European Wasp along the Adelaide Hills, any existing butterfly colonies in this area are
likely to be terminally affected by this very efficient predator wasp.
-
Conservation Strategy
-
The butterfly requires broad acre pristine habitat for survival. Colonies of early stages
need to be left alone. Butterfly collectors should target flying adults, preferably males.
If collectors must have perfect specimens, then only a few mature larvae minimally
collected from the hostplant at night during the spring and early summer months should occur.
Byres need to be left unbroken.




