-
Larval Food Host
-
In South Australia, larvae have only been found on the mistletoes
Amyema miquelii (box mistletoe) and
A. pendula pendula (drooping mistletoe) (Loranthaceae).
The larvae eat the flower buds, flowers, leaves and soft stem parts of the mistletoe,
but are particularly fond of the flower buds. The mistletoes usually associate with
Eucalyptus, although A. pendula will also parasitise Acacia melanoxylon,
particularly in the Lower Southeast Region of the state (where the butterfly has yet to be found).
In the eastern states other mistletoes are also utilised as hostplants, and those
that can be found in SA comprise Amyema miraculosa (fleshy mistletoe)
and A. quandang (grey mistletoe).
-
Larval Attendant Ant
-
Larvae found along the Mt Lofty Ranges (that generally equates to the
distribution of form genua) are attended by numerous large
sugar-ants Camponotus consobrinus.
Elsewhere in South Australia (that generally equates to the distribution
of form splendida), larvae are attended by numerous, very large
sugar-ants Camponotus nigriceps.
Both these ants prefer heavy loam soils, although the latter also build
nests in tree hollows above the ground if the soil is sandy. A similar
ant attendance preference is also apparently found elsewhere in Australia,
with butterflies that occur in coastal and moist areas being mostly
attended by C. consobrinus, whereas butterflies found in inland
areas being usually attended by C. nigriceps. The very large and
aggressive C. intrepidus is also an attendant ant in the eastern
states, but this ant is not known to occur in South Australia.
The ants are nocturnal, large, only moderately ferocious (but without a sting),
although C. nigriceps can sometimes be aggressive. They have formed a
symbiotic relationship with the butterfly, whereby the ants provide protection
to the early stages of the butterfly from predators and parasitoids, and in
return the ants receive sugary secretions from the median dorsal secretionary
organ (honey gland or Newcomer's organ) situated at the posterior end of the
larvae. Adult ants are able to live off these secretions. Unlike the situation
with Ogyris subterrestris in which
adult butterflies are often killed by the ants on contact, Ogyris genoveva
is not attacked unless it remains within the ant chamber after emergence from
the pupa (if deformed or blocked in by other pupae). However, the butterflies
often become agitated in the presence of the ants and will distance themselves
from any unnecessary attention. (It only requires a bight from these large ants
to inflict a mortal injury. )
The ants essentially farm the larvae. They build separate chambers away from the ant
nest to house the larvae. The ant chambers or galleries are specially constructed
below ground around the base of the host tree by the attendant ants for sheltering
the early stages of this butterfly. There can be multiple entrances, but they are
always narrow and easily guarded by the large Camponotus ants. These chambers
can be quite extensive, circling the base of the tree and extending along parts of
the roots. The gallery space between the tree and the ground is usually not much wider
than the mature larvae or pupae. Sometimes larvae will use hollowed out branches that
are lying on the ground next to the base of the host tree. Camponotus nigriceps
will more often use hollows within the host tree for use as a gallery. These ant
chambers are usually widely separate from the actual ants' nests (or ant brood), but
sometimes butterfly larvae will occur near the nest, but in this situation the ants
construct a separate chamber adjacent to the nest to house these larvae which is
separate from the ant brood.
The ants provide 24-hour protection, and when the larvae leave the chamber at night to
feed on the mistletoe, they are guarded non-stop until they return to the chamber again
before dawn. The ants also keep the chamber clean of larvae frass (excrement) and moult
skins. Studies have shown the incidence of larvae parasitisation, probably the main way
the larvae are predated, are about 30% with C. consobrinus, but can be reduced to
as low as 5% by the extremely aggressive C. intrepidus (Eastwood 1997).
C. consobrinus
is a common, large, orange and black coloured ant, that frequently enters houses at night
in urban areas, where it is known as the sugar-ant. It is totally harmless to humans,
unless one is allergic to formic acid. It should not be confused with the large stinging
and dangerous
bull-ants, inch-ants or jumping ants.
The latter group have highly visible, very large and long jaws projecting down from the head.
In C. consobrinus the jaws are not readily visible. C. nigriceps is common in
pastoral areas, and is a large, yellow and black coloured ant. It is strictly nocturnal.
-
Eggs
-
Large, hemispherical shape, strongly flattened top and bottom, the sides are very finely
reticulated in a hexagonal pattern, with each reticulation intersection having a small
raised blunt projection, the facets on the top of the egg are smaller and these continue
to the small-depressed micropylar area at the apex of the egg. The basal half of the egg
is greyish white, the top half is mostly dark brown with a greyish white ring near the
outer dorsal edge, and the micropylar area is greyish white. The eggs are typical for the
species-group. Some eggs may be a washed out grey or white colour, and near larval emergence
the egg can sometimes turn pale grey. The larva emerges from the egg at night by eating out
the dark brown coloured (central) dorsal area. (The parasitic wasps, one per egg, emerge
from the egg by eating out a smaller circular area in the dorso-lateral region.) Eggs hatch
after 6 days in summer. (The parasites emerge after about 20 days).
Using their sensory organs, the females actively seek out mistletoe hostplants that already
support existing early stage colonies of the butterfly and their Camponotus attendant
ants. Their antennae and palps probably locate the ants and mistletoe, but their ovipositor
also seems to be involved in the process of locating the correct place to lay eggs. In the
Adelaide Hills the females are often seen on the mistletoes within a known colony, walking
all over them, testing the stems with their ovipositor without actually laying eggs. Using
their sensory organs they eventually make their way down the trunk of the mistletoe host
tree following the larva/ant trails (between the ant chambers at the base of the tree and
the mistletoe high up in the tree), to locate the attendant ant chambers, and then they
will start to lay eggs in batches. These batches can vary from just a few eggs up to large
collections of eggs, and 46 eggs have been recorded in one large batch. The eggs are usually
laid adjacent to each other to a depth of one egg, but in large batches they can be stacked
upon each other two or three deep. The batches are laid beneath bark or within cracks of the
tree trunk, usually within two metres of the ant chambers, which may be within a hollow in
the tree or in the ground at the base of the tree. Older females will also sometimes lay
one or two exposed eggs on the larvae/ant trails on the host tree. The females will only
lay where there is a strong residual smell of the attendant ants, and also prefer hostplant
which is already being utilised by the early stages of the butterfly. They prefer host trees
in which the mistletoe is not too high off the ground, for obvious beneficial reasons for
the larvae. The C. consobrinus ants prefer trees which have some loose vegetal debris
about their bases.
In major colonies, there is a large incidence of egg parasitisation, as the eggs are
unprotected by the ants, and the parasitoid wasp is too small for the ants to regulate.
In an ideal environment the egg parasitoid probably keeps the colony to reasonable size
limits, as otherwise the larvae would likely build up to enormously large numbers that
would quickly eat all its mistletoe hostplant and result in the demise of the colony.
The downside is that if the colony is stressed, the wasps could actually cause the demise
of the colony. During the 2003 flight season (a drought season), a single large colony
tree in the Adelaide Hills was examined late in the season, and from 265 eggs examined
and it was found that 147 eggs had hatched normally, while 100 eggs had been parasitised
and 18 eggs had remained unhatched.
-
Larvae
-
The first instar is long onisciform shaped, weakly scalloped laterally, initially purplish
brown coloured with a greyish white perimeter, and a broad broken greyish white longitudinal
dorsal line that terminates at abdominal segment 6. After eating the hostplant, the pale
areas turn pale greyish yellow, and the pale broken dorsal line becomes a solid line.
The anterior and posterior ends are dark brown. The posterior dorso-lateral organs are not
developed. The head is large, smooth, yellowish brown, hidden beneath the body. There are
long peripheral and dorsal setae (hairs), which are longer anteriorly and posteriorly.
The dorsal setae terminate at abdominal segment 6. The peripheral setae are mostly white,
with a few black setae at the posterior end. Dorsal setae occur in two pairs, one black
pair being long and recurved, the other pair being white coloured, short, recumbent and
directed to the rear, except on the sixth abdominal segment where both pairs of setae are
black, long and recurved.
The newly hatched first instars are believed to make their way down the trunks from the
egg batches to the base of the trees to the ant chambers where they are probably rounded
up by the ants and herded in with the rest of the larvae. The young larvae are very mobile
due to well-developed thoracic legs, and are quite capable of climbing the host tree with
ease every evening. (The first instar is about 2 mm long when it hatches from the egg).
However, as the first instars do not possess posterior dorso-lateral organs the ants do not
show as much interest in them as they do with older larvae that possess these organs. First
instar larvae eat by scouring the leaf or bud surface, but older larvae eat from the leaf
edge or devour the entire bud.
Intermediate instars gradually lose the long setae, and they are also strongly scalloped
laterally. From the second instar, the dorso-lateral organs are prominent at the posterior
end. The second, third and fourth instars have a similar colour to the fifth instar, but
are mostly purplish brown coloured with the pale yellow markings being less obvious.
The butterfly tends to occur in colonies, of variable size, with some old colonies that have
not been interfered with, reaching half a hectare or more in areal extent, with most of the
mistletoe bearing trees in the area having early stages of the butterfly. Due to past
clearing and fragmentation of habitat, it can be many kilometres before another colony is
encountered. However, females will disperse from the colonies and on occasions, solitary
trees are found with early stages of the butterfly. Some large individual trees in a colony
can harbour enormous groups of larvae and pupae, and there are eastern states records of 200
or more individuals. In South Australia the documented numbers are less than 100, usually
much less. Colonies are prone to communal diseases like bacterial and virus infections,
which are transmitted by all the chamber occupants, including the ants.
The final instar grows about 30-33 mm long, but larvae distended by parasites may reach 40 mm
in length. They are mostly dull purplish brown coloured, with pale yellow dorsal chevron
markings and other indistinct subdorsal markings, the chevrons on abdominal segment 6 are
joined together to form a yellow triangular mark, the lateral and anterior extremities of
the larvae are pale yellow, the prothoracic plate is dark grey with a yellow longitudinal
dividing line, the anal plate is dark brown and yellow with a black longitudinal dividing
line, and the spiracles are black. It is long onisciform shaped, with a thoracic dorsal
furrow, the lateral edges are scalloped, the anterior and posterior areas are flattened,
there are some short peripheral bristly hairs, and there are some short dorsal bristles
on thoracic segment 3 and abdominal segments 1-6. The body is covered in dark coloured,
minute secondary setae, which consist of a short simple pointed shaft set on a protuberant,
angular and ridged base. The secondary setae impart a scabrous appearance to the larvae.
The posterior dorso-lateral organs are well developed. The head is small, smooth, brown,
hidden beneath the body.
Larvae feed only during the night, accompanied by large numbers of attendant ants. The ants
usually become active before the larvae and numbers of them head towards the mistletoe to
make sure the path is clear of any obvious larva predators, like spiders. The larvae begin
to emerge from the ant chamber shortly after dusk. The early instar larvae are often the
first to emerge from the chambers, and quickly head off for the mistletoe, which can be up
to 20 m away, high up in the tree canopy (although it is usually much less). Older instars
are often more timid and slower to get moving, as the least disturbance can disorientate
them. Eventually, a large procession of larvae and protective ants head off for the mistletoe.
Larvae leave a silken trial as they climb the host tree to the mistletoe, which is discernible
under certain lighting conditions. They always follow the same trail every night. Larvae in
attendance by ants continually extrude their white coloured, extendable posterior lateral
(tentacular) organs, which emit pheromone chemicals that convey an attentive warning signal
to the attendant ants (i.e. "make sure you are looking out for me" ). The ants are often seen
to ride on the backs of the large larvae as they move about on the host tree. Presence of
larvae on a hostplant is readily discernible by larval eat marks on the mistletoe leaves,
and they feed independently on the mistletoe (not gregariously). Larvae usually feed on
warm nights, and return to the ant chamber before dawn.
During the day the larvae hide from predators by remaining in the ant chambers. The ants
actively guard the entrances to the chambers. Larval excrement (frass) and the discarded
skins from the moulting larvae within the chambers are removed by the ants. In South
Australia, larvae attended by the less aggressive C. consobrinus have a high
incidence of larval parasitisation by small braconid wasps (Apanteles sp). Some 30
or more wasp larvae will parasitise within a single large O. genoveva larva, and all
will exit at the same time then collect beneath the butterfly larva to pupate, creating a
white silken mass of tiny wasp pupa-cases. The exiting of the wasp larvae paralyses the
butterfly larva, but ants continue to attend the stricken larva, which will take a few days
to eventually die. Interestingly, the ants do not attack either the exiting wasp larvae or
the adult parasitoid wasps, so it is likely the latter two have pheromones that mimic those
of the butterfly larvae and attendant ants.
Larvae seem to require six instars to fully develop, and records of seven instars are known.
The larva period is variable, depending on the location, time of the year, and the condition
of the hostplant. During winter, larval development is very slow, and essentially ceases once
the average temperature drops below 15 C. Over the warmer months the colonies usually contain
larvae at all stages of development although mature larvae occurring outside of the main
flight period are likely to be parasitised.
-
Pupae
-
Long cylindrical, rounded anteriorly and posteriorly, about 18-22 mm long, dark brown
to blackish brown, with a few very short lateral hairs on the abdomen. The surface of
the pupa bears a very fine reticulated pattern (similar to the eggs) that produces a
scabrous appearance. The larvae are strongly attached to the silked substrate by anal
hooks and a central girdle, usually to any non-earthen object or surface below ground
level within the ant chamber, such as the bark of the host tree or rock or stick or
vegetation caught up within the construction of the chamber, or if the colony is large,
to the roots of the host tree. Sometimes the larvae will pupate within white ant hollowed
stumps or fallen boughs next to the base of the host tree, or in the case of
Camponotus nigriceps in hollowed boughs or cavities higher up in the tree trunk.
Occasionally the larvae will pupate beneath loose bark near the entrance to the chamber
either on the host tree or on the ground, but often these larvae are parasitised. The
larvae pupate gregariously together, and sometimes very large numbers of pupae can occur.
The butterflies usually emerge during the morning, and expand and harden their wings either
within the chamber entrance if it is large enough, or on the mistletoe host tree up to 2 m
above the chamber entrance.The ants do not normally attack the butterflies during this stage,
although the ants will eventually destroy butterflies that emerge deformed or are unable to
exit the chamber for any reason. The empty pupa shells are also destroyed by the ants, or if
they do not do it quickly enough then another pupating larva may remove it to make room for
its own pupation.
Butterflies emerging from the pupae are not covered in large quantities of fluffy grey 'down',
which is known to occur in many other Ogyris. (This down helps protect the butterfly
from being attacked by over-zealous attendant ants, or even other small predators, while the
butterfly is expanding and hardening its wings in readiness for first flight.) Since
O. genoveva often expands its wings within the presence of the protective attendant
ants (which will not bother the butterfly unless either become very agitated), there is
apparently no need for the predator-repellent down.
The pupae are not known to audibly stridulate, although interestingly, the prepupae can
stridulate, making a series of audible clicks, which are believed used as a means of
communicating with the attendant ants.
The pupa period is highly variable, depending on the location and time of year. In the South
Mt Lofty Range the pupa period is about 34-35 days in late spring, reducing to about 14-24
days in mid-summer and early autumn. In the Olary Ranges the pupa period is about 33-38 days
in early spring.
-
Flight Period in South Australia
-
The butterfly flies throughout the warmer months with records from September to early June.
In hot northern areas there seems to be 3 flights starting in September-October, where the
butterfly tends to start flying earlier and finish later in the season compared to southern
areas. The main flight in cool, southern areas occurs in January and February to usually
coincide with the flowering of the mistletoe hostplants, which are used for nectaring
purposes by the adults. In captivity, a brood can be completed in 12 weeks over summer-early
autumn in the South Mt Lofty Range, and this could theoretically account for a much smaller
secondary autumn flight that is sometimes witnessed in certain populations, but as with
most colonistic Lycaenids there is staggered larva development which could also lead to an
autumn flight. In the South Mt Lofty Range most of the larvae resulting from eggs laid
during summer continue to develop over winter to form next seasons flight. The butterfly
over-winters as larvae.

-
Distribution
-
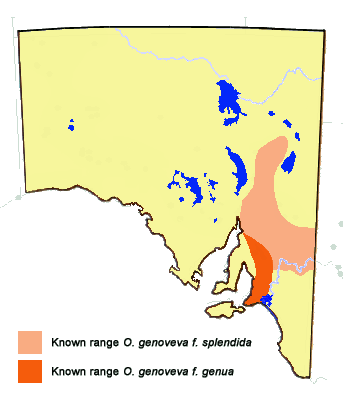
The butterfly is presently known from the Mt Lofty, Flinders and Olary Ranges, and the
Riverland areas of the state. Its hostplants are common in the Southeast Region, but
the soil type is mostly white sand or ephemeral wetland in which the likely attendant
C. consobrinus ants are unable to live. The butterfly also occurs in Vic, NSW
and southeast Qld.
The habitat between the Flinders Ranges and Broken Hill/Mildura where the butterfly
occurs is similar to the habitat between the Flinders Range and the Nullarbor Plain
but the butterfly has yet to be seen west of the Flinders Ranges, even though many
concerted efforts have been made to try and find it. The presumed absence of
O. genoveva from that area is puzzling suggesting that either this species
is a recent immigrant to SA from the eastern states and has yet to breach this
western area of SA or that the region west of the Flinders Ranges has suffered a
previous relatively recent climatic event (global warming) that has eliminated the
species from the area.
-
Habitat
-
The butterfly can occur in a variety of habitats, from moist cool temperate to hot and
dry semi-arid conditions. It is not found in arid or alpine areas. Limiting factors to
its range appear to be a requirement for an average annual minimum temperature between
about 6 and 12 C. It is most frequently found in open (Eucalyptus) woodland, and
will sometimes briefly venture into the outer urban environment. The obligate attendant
ants are widespread.
-
Conservation Status in South Australia
-
The butterfly is usually rare in its occurrence, but widely distributed. It can sometimes
be locally common where it is well established in undisturbed colonies.
-
Threats
-
The habitat along the Mt Lofty Range and in the Riverland has been extensively fragmented
and destroyed by past clearing efforts. This habitat continues to be destroyed for small
acre farms and urban expansion. Remaining habitat in the Mt Lofty Ranges is also subject
to frequent bush fires. In urban areas its mistletoe hostplants are usually considered
pests and often removed from their hosts. Elsewhere, the butterfly is not generally under
threat except where the habitat is adjacent to farming communities and affected by poison
spray drift during crop spraying activities, especially when the poison is applied by
aerial means. Parts of its range within the Southern Flinders and Olary Ranges are within
the prime breeding grounds for the
plague locust and
the butterfly would be decimated by any toxic spray programs adopted by the Locust Control
Board (as happened in year 2000). Burrowing animals such as rabbits and echidnas have the
potential to destroy larva colonies at the base of the host trees. Stressed colonies may
also be terminally weakened by the collective actions of parasitoid wasps that attack both
eggs and larvae, such that adult butterfly numbers become too low to effectively maintain
the continuance of the colony.
-
Conservation Strategy
-
Spraying activity should be carried out judiciously. In urban areas a public education
process is required for the Amyema mistletoe. Healthy trees are able to support
this mistletoe, and there is usually a self-induced balance between the tree host and
the mistletoe. If there appears to be an imbalance then it should not be a problem to
thin out some of the mistletoe. An active Ogyris colony will help keep the
mistletoe pruned. Ideally, every landowner should carry out their own property survey
for this magnificent butterfly, by checking for larvae on mistletoe bearing host trees
during a warm evening. If the larvae are present, then measures can be put in place by
the landowner to not disturb the colony. Actions that can destroy the larvae colony
include, digging around the base of the host tree, removing the host tree or mistletoe,
removing debris from around the base of the host tree, insecticide spraying and burning
off. Digging into the colony exposes the early stage larvae and pupae to predators,
particularly parasitoids and other ants such as Iridomyrmex spp, which will quickly
destroy the colony. The attendant Camponotus ants cannot fully protect the
early stages once the colony chamber has been opened.







